Coweeta LTER History
Funding for the Coweeta LTER program (1980-2020) has been terminated by NSF. The USDA Forest Service Coweeta Hydrologic Laboratory, however, continues to operate normally (https://www.srs.fs.usda.gov/coweeta/ ). This static website briefly describes the Coweeta LTER research program (below). Other pages on this site provide alphabetical and chronological bibliographies of all publications through 2020, a set of highlighted publications, experimental basin research history through 2020, and a list of past LTER researchers. Coweeta LTER data and publications can also be accessed through the NSF LTER website and the Coweeta Hydrologic Laboratory website. 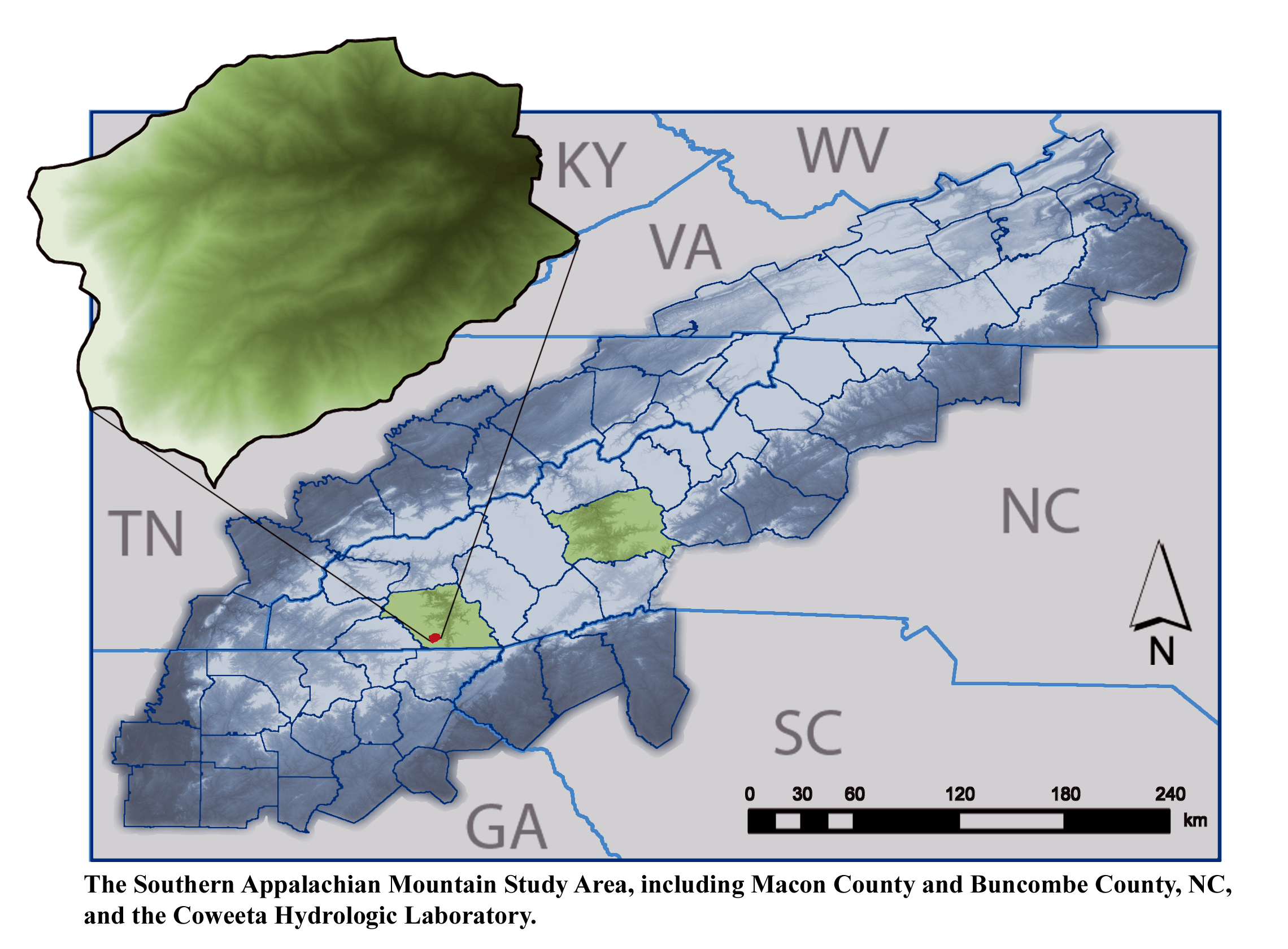
Program History
Long-term ecological research at Coweeta has been the product of a series of collaborative agreements between the University of Georgia Research Foundation in Athens, Georgia, and the USDA Forest Service Coweeta Hydrologic Laboratory in Macon County, North Carolina, with many other cooperating Universities. These research agreements preceded NSF funding of the Coweeta LTER (CWT) in 1980 as one of six original sites in the US LTER Network. CWT field operations are based out of the Coweeta Hydrologic Laboratory. CWT has been dedicated to developing process understanding of ecological dynamics in montane deciduous forests of the southern Appalachians. An overarching question has guided CWT research:
How do natural disturbances, topography, climate variability, and human activities interact with ecological processes and ecosystem states in temperate montane deciduous forests?
The Forest Service Hydrologic Laboratory itself was established in 1934, and much of the infrastructure was built by the Civilian Conservation Corps. Research at the Laboratory was originally focused on questions in forest hydrology, for which the site was world-renowned. In 1968, the Laboratory’s research mission was expanded to include forest and stream ecology, including factors regulating biogeochemical cycling and exports, organic matter production and turnover, and ecosystem responses to climate change. The original partnership between University of Georgia and the Coweeta Hydrologic Laboratory was signed in 1968 and since then has been the basis for multiple collaborations in national and international research programs. These partnerships have included the International Biological Program, the TIE Experimental Ecological Reserves Program, the International Hydrologic Decade, and the UNESCO Man and the Biosphere Program. Most significant, however, has been collaboration on the NSF-funded Coweeta LTER (CWT).
In 1994, the geographic and conceptual scope of CWT research expanded to include fundamental socio-ecological questions relevant to the larger Appalachian region, specifically within the Upper Little Tennessee and Upper French Broad River Basins. Southern Appalachian forests are among the most biodiverse and productive in the temperate world, but they are also experiencing some of the most rapid exurban growth in the continental U.S. Consequently, Coweeta LTER research was organized to be simultaneously place-based, cross-scale, long-term, and comparative, reflecting how the program has adapted ecological theory and practice to the present and future needs of society in the southeastern United States. This mixed research scale allowed for long-term detailed mechanistic study of relatively undisturbed systems at the core, and equally-relevant studies of the interactions of ecological and social processes involved with human activities that dominate ecosystems at regional scales. Universities participating in CWT have included Virginia Tech University, Duke University, University of Wisconsin, University of Minnesota, Mars Hill College, University of Illinois, University of North Carolina, North Carolina State University, SUNY-Buffalo State, Indiana University, Appalachian State University, and UNC-Charlotte. NSF funded CWT in cycles, and research within each cycle focused on different aspects of the core question (Figure 1). In 2016, NSF decided to discontinue funding of the Coweeta LTER, and NSF-funded research projects and operations were completed or phased out from 2016-2020. Collaborative ecosystem research will continue at the site in collaboration with the USDA Forest Service Hydrologic Laboratory, but without the funding and structure of the LTER program.
Ecosystem Overview and History
Appalachian ecosystems are unique in their antiquity, temperate biodiversity, high ecosystem productivity, and steep natural environmental gradients that have been modified by human disturbance. The unglaciated Southern Appalachian Mountains acted as a habitat refuge for many species during continental glaciation and the interaction of the local climate with the rugged topography creates a rich diversity in habitats. The region’s habitat diversity has produced high biodiversity in plants, fish, and salamanders. The Southern Appalachian ecosystem has been variously and highly disturbed since European settlement (Figure 1), yet appears to exhibit high resilience relative to many ecosystems.
During Native American settlement of the region, mixed deciduous forests covered the hillslopes, but the understory was regularly burned to improve wildlife habitat. Row crop agriculture was practiced in the valleys. In the early 1800s, European settlers forcibly relocated most of the Cherokee inhabitants of the region to Oklahoma, making way for the European immigrants to clear the forests and attempt agriculture on both hillslopes and valleys. The impracticality of hillslope agriculture was soon apparent from large-scale erosion and valley sedimentation, and the settlers focused on valley agriculture, allowing the hillslope forests to regenerate naturally beginning in the late 1800s and early 1900s.
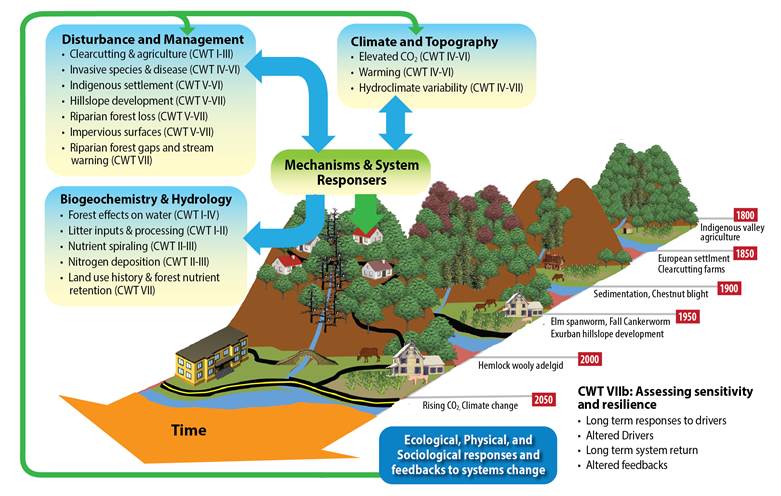 Figure 1. Conceptual framework guiding CWT LTER research. CWT has sought to understand how historic and current human activities affect populations, biodiversity, biogeochemical cycling and other ecosystem processes and patterns. Early efforts determined the effects of forest clearing and agriculture on biogeochemistry, productivity, and hydrology (CWT I through III) (e.g. Swank and Webster 2014). Continuing research expanded our investigations of anthropogenic disturbances, to span the Holocene (CWT IV, V). These long-term data sets have allowed us to analyze ecosystem response rates to disturbances and climate change and to discover surprises in the records (CWT VI). All of our work builds on the long-term observations of system behavior, but our research portfolio also includes short-term mechanistic and gradient studies that explicitly recognize the dominance of human activities over many states and processes, and the corresponding social and economic drivers (CWT V through VII).
Figure 1. Conceptual framework guiding CWT LTER research. CWT has sought to understand how historic and current human activities affect populations, biodiversity, biogeochemical cycling and other ecosystem processes and patterns. Early efforts determined the effects of forest clearing and agriculture on biogeochemistry, productivity, and hydrology (CWT I through III) (e.g. Swank and Webster 2014). Continuing research expanded our investigations of anthropogenic disturbances, to span the Holocene (CWT IV, V). These long-term data sets have allowed us to analyze ecosystem response rates to disturbances and climate change and to discover surprises in the records (CWT VI). All of our work builds on the long-term observations of system behavior, but our research portfolio also includes short-term mechanistic and gradient studies that explicitly recognize the dominance of human activities over many states and processes, and the corresponding social and economic drivers (CWT V through VII).
As the climate in southern Appalachia changes, the distribution and behavior of mesophytic plants and animals are being altered and ecosystem function is likely to change. Coweeta LTER research sought to understand how climate change and development affect biodiversity at local to regional scales and the implications for ecosystem processes.
Select Highlights of CWT Research (see bibliography of signature publications)
Watershed 7 (WS7) Basin-scale clearcut and natural regeneration experiment.
In the 1970s, the Forest Service clear-cut Watershed 7 without riparian buffers using the Best Management Practices of the time and allowed the forest to regenerate naturally. This manipulation was conducted to observe the magnitude of ecosystem response and the rate of recovery to pre-disturbance conditions. Continued long-term observations demonstrated that this view of resistance and resilience was too simplistic. Forest disturbance triggered a chain of ecological dynamics that are still evolving after 40 years
After harvest, microbial nitrogen mineralization continued, but plant uptake was diminished, so streams experienced a short-term pulse in dissolved inorganic nitrogen as expected. Similarly, reduced interception and transpiration caused streamflows to increase for four years. Afterwards, the system appeared to be returning to pre-disturbance conditions according to ecosystem resilience concepts. However, unanticipated changes in forest composition, specifically early-stand establishment of a large component of nitrogen-fixing black locust trees, caused stream DIN levels to increase again and remain elevated for over at least 40 years. Eventually, forest composition changes also led to reductions in streamflow. These long-term observations of streamflows, stream dissolved inorganic nitrogen (DIN) concentrations, stream DIN exports, and stand composition substantially advanced our understanding of forest ecosystem dynamics. Furthermore, they demonstrated that long-term observational data can reveal ecosystem complexities and surprises, generating new hypotheses and motivating new research. For example, the long-term DIN response to black locust establishment motivated new research into the timescales and ecosystem effects of enhanced biological nitrogen fixation following disturbance.
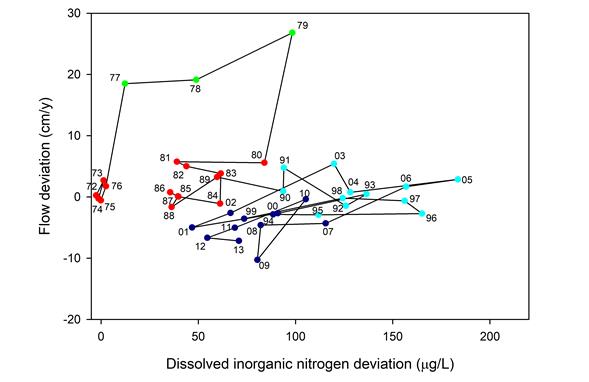 Figure 2. Progression of annual DIN and discharge deviation between Coweeta Watersheds 7 (treatment) and 2 (reference) from 1973–2013. The discharge deviation is the difference between the observed WS7 flow and that predicted from WS2 based on the pre-treatment model. The DIN deviation is WS7 DIN – WS2 DIN. Each point is marked by the last two digits of the year. Symbol colors denote separate clusters identified by k-means clustering by year. Green points encompass the initial watershed response (1977–1979) featuring large increases in water yield and moderate nitrogen release after harvest. Red points encompass the pre-harvest period (1972–1976), but also a period when the watershed seemed to be returning to pre-harvest conditions (1980–1988) after the initial response. Light blue points encompass the period of high DIN concentrations (1990–1998) due to nitrogen fixation following the emergence of Robinia as a dominant forest tree. Dark blue points encompass the most recent period (1999–2013) featuring lower streamflows and DIN. These latter two periods suggest interactions between droughts and DIN. The data do not yet indicate a move back to the initial condition
Figure 2. Progression of annual DIN and discharge deviation between Coweeta Watersheds 7 (treatment) and 2 (reference) from 1973–2013. The discharge deviation is the difference between the observed WS7 flow and that predicted from WS2 based on the pre-treatment model. The DIN deviation is WS7 DIN – WS2 DIN. Each point is marked by the last two digits of the year. Symbol colors denote separate clusters identified by k-means clustering by year. Green points encompass the initial watershed response (1977–1979) featuring large increases in water yield and moderate nitrogen release after harvest. Red points encompass the pre-harvest period (1972–1976), but also a period when the watershed seemed to be returning to pre-harvest conditions (1980–1988) after the initial response. Light blue points encompass the period of high DIN concentrations (1990–1998) due to nitrogen fixation following the emergence of Robinia as a dominant forest tree. Dark blue points encompass the most recent period (1999–2013) featuring lower streamflows and DIN. These latter two periods suggest interactions between droughts and DIN. The data do not yet indicate a move back to the initial condition
Climate and Forest Change
CWT’s long-term climate records indicate that spring, summer, and annual air temperatures are increasing, with more very hot summer days (Oishi 2018). Precipitation variability is increasing (Burt et al. 2018, Laseter 2012). Spring leaf-out is occurring earlier than in previous decades due to warmer spring temperatures. Leaf senescence arrives earlier following growing season droughts at rates that depend on elevation (Hwang et al. 2011a and b, 2012, 2014). Hydrologic modeling indicates that current trends will lead to reduced soil moisture in summer and fall at both low and high elevations, uncertain changes in summer streamflow, and increased streamflow in winter (Wu et al. 2011, 2012). These temperature-moisture interactions have impacts that vary widely between tree species and size classes.
The high habitat and biotic diversity of the region creates a high-dimensional system controlling forest response to climate change. Air temperature-soil moisture interactions have impacts that vary widely between tree species and size classes. Clark et al. (2013) developed the dynamic inverse prediction framework for quantifying interactions among drivers and responses in complex high-dimensional systems. Clark et al. (2013) used this framework to show that the most important variables for forest change at individual scales can be extended to explain patterns at continental scales (Zhu et al. 2012, Zhu et al. 2014). Forest diversity in this ecosystem of intense competition is higher than can be explained by ecological theory. Models predict low diversity, as the best competitors should drive all others to extinction. Clark’s work revealed that high biodiversity can be maintained by interactions and indirect effects that involve temperature, drought, and local moisture availability (Clark 2010, Clark et al. 2010, 2011b, 2012b, Dietze and Clark 2008, Uriarte et al. 2012). Individual trees respond most like other trees of the same species, a mechanism that concentrates competition within species (Clark 2010, Clark et al. 2010, Clark et al. 2012a, b, c). Interactions involving climate and competition also control understory plant abundance, reproduction and growth across gradients (Warren 2009, Warren and Bradford 2010). Interactions between native and nonnative understory species vary systematically with climate (Fraterrigo et al. 2014).
The most significant ecological alteration to southern Appalachian forests, the loss of the dominant chestnut tree to the introduced chesnut blight, happened long before the CWT LTER was established, but long-term vegetation plots created by the USFS have allowed CWT to track large changes in forest composition over time. Climate trends would suggest the forest should have shifted towards an oak-hickory (OH) forest (McEwan et al. 2011), in agreement with vegetation models (Iverson et al. 2004). However, our long-term data indicate proliferation of mesophytic species such as red maple and tulip poplar (MP) (Elliott and Vose 2012, McEwan et al. 2011, Caldwell et al. 2016), henceforth mesophication (sensu Nowacki and Abrams 2008). The reason for the lack of alignment between long-term records and climate models is unclear. While oaks are dominant in the region, their regeneration over the last century is lower than that of red maple and other mesophytic species (McEwan et al. 2011). Fire exclusion could explain recent decline in oaks (Abrams 1992, 1998, 2003). In the Coweeta Basin, tree recruitment has also been suppressed by the expansion of rhododendron, a consequence of American 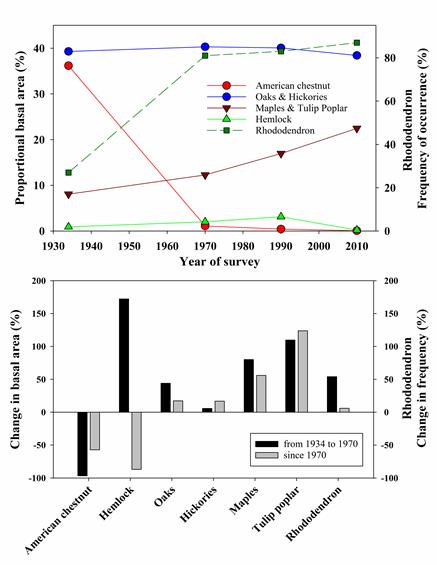 chestnut and hemlock loss. CWT research shows that the dense rhododendron subcanopy strongly attenuates light (Clinton 2003), suppresses herbaceous plants and tree seedlings (Clinton et al. 1994, Beckage et al. 2000, HilleRisLambers and Clark 2003), and reduces nitrogen availability in the soil mineral and organic horizons (Wurzburger and Hendrick 2007, 2009). Our long-term forest demography data suggest that a chain of disturbances -- starting with widespread logging in the 19th century, the loss of American chestnut, and finally the hemlock woolly adelgid invasion -- led to a cascade of overstory, midstory, and soil responses that permitted an expansion of rhododendron. Therefore, the response of future forests to hydroclimatic extremes may depend on these tree-shrub interactions that manifest through disturbance legacies and indirect effects.
chestnut and hemlock loss. CWT research shows that the dense rhododendron subcanopy strongly attenuates light (Clinton 2003), suppresses herbaceous plants and tree seedlings (Clinton et al. 1994, Beckage et al. 2000, HilleRisLambers and Clark 2003), and reduces nitrogen availability in the soil mineral and organic horizons (Wurzburger and Hendrick 2007, 2009). Our long-term forest demography data suggest that a chain of disturbances -- starting with widespread logging in the 19th century, the loss of American chestnut, and finally the hemlock woolly adelgid invasion -- led to a cascade of overstory, midstory, and soil responses that permitted an expansion of rhododendron. Therefore, the response of future forests to hydroclimatic extremes may depend on these tree-shrub interactions that manifest through disturbance legacies and indirect effects.
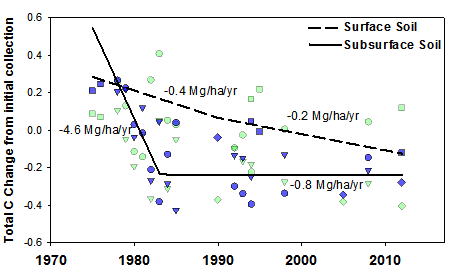
Following the decline of American chestnut, Eastern hemlock (Tsuga canadensis L.) expanded substantially in riparian areas (Woods and Shanks 1959, Elliott and Swank 2008), but declined precipitously at the turn of the 21st century as a result of the hemlock woolly adelgid (Adelges tsugae Annand) invasion). Our work has revealed cascading ecosystem effects of hemlock loss. Hemlock mortality reduced soil carbon pools and fluxes in the short-term (Nuckolls et al. 2009, Ford et al. 2012); caused initial reductions in forest water use followed by increases (Ford and Vose 2007, Brantley et al. 2013, 2014); reduced decomposition (Ball et al. 2008); and caused changes to nutrient cycling that were dependent on the presence of rhododendron (Knoepp et al. 2011, Block et al. 2012, 2013). Stream responses to hemlock loss are manifested through a cascade of forest species changes, which affect the quantity and quality of particulate organic carbon (POC, Wallace et al. 1997 Wallace et al. 2015), hydrology and stream chemistry (Knoepp et al. 2014, Qualls et al. 2014, Meyer et al. 2014), and stream production (Cross et al. 2007). The mortality of hemlock now appears to favor the growth and density of rhododendron (Rhododendron maximum L.) over other woody species (Ford et al. 2012, Elliott and Miniat 2018, Dharmadi et al. 2019). Increases in riparian rhododendron growth and density after hemlock loss decreased microbial and invertebrate diversity and litter decomposition rates (Kominoski et al. 2009, Kominoski and Pringle 2009). Light incident on streams increased following hemlock loss, increasing trophic processing (Webster et al. 2012a) and shifting stream metabolism towards greater heterotrophy (Northington et al. 2013). While the loss of riparian hemlock has had direct effects on stream ecosystem woody debris, light transmittance, and temperature variation (Webster et al. 2012), the longer-term effects of hemlock loss on both terrestrial and aquatic ecosystems will likely depend on the interactions and indirect effects that manifest from the expansion of rhododendron. Together, these changes in forest species composition have coincided with declines in deep soil carbon (Fig. 3, Knoepp et al. 2014) and water yield (Caldwell et al. 2016).
 Land use change and climate change also affect biodiversity of higher trophic levels. Increasing mountainside development has reduced habitat for interior-forest neotropical migrant bird species (Lumpkin et al. 2012, Lumpkin and Pearson 2013). These species appear vulnerable to warming and increased nest predation, open forest canopies, and increases in predatory mammals common in residential areas. Community composition of herb species depends on interactions involving elevation, competition with other herbaceous species, the presence of ants, and land use legacies (Jackson et al. 2012).
Land use change and climate change also affect biodiversity of higher trophic levels. Increasing mountainside development has reduced habitat for interior-forest neotropical migrant bird species (Lumpkin et al. 2012, Lumpkin and Pearson 2013). These species appear vulnerable to warming and increased nest predation, open forest canopies, and increases in predatory mammals common in residential areas. Community composition of herb species depends on interactions involving elevation, competition with other herbaceous species, the presence of ants, and land use legacies (Jackson et al. 2012).
Stream Ecology
Stream ecology at Coweeta focused on the connections between streams and their watersheds. CWT researchers studied how differences in riparian and hillslope land cover, soils, and infrastructure alter connectivity of water, solutes, energy, and organic matter from hillslope to stream to river. These studies included observational studies of how streams are modified by forest practices and changing land use, experimental studies primarily addressing stream-watershed connections, and cross-site syntheses to evaluate generalities about how streams reflect watershed conditions. The original WS7 experiment raised questions about how changes in incident light, organic matter input, streamflows, and DIN concentrations affected primary productivity, aquatic assemblages, and biogeochemical cycling in streams. These questions were further explored through experimental manipulations of other streams. A 13-year manipulation of litter inputs (first exclusion, latter addition) demonstrated that biomass and of functional feeding groups were tightly linked to litter inputs, except in rockface habitats with little exclusion project. A streamflow reduction manipulation showed that litter availability as a basal resource was strongly linked to streamflows. A stream nutrient addition experiment showed that elevated nutrient concentrations stimulate leaf litter breakdown and accelerate stream carbon loss. The cross-site LINX experiment showed that small streams retain a large fraction of inorganic N inputs and play a disproportionately large role in stream network nutrient cycling.
The first stream studies examined effects of sediment from logging on fish and stream insects (Tebo 1955, 1956) and also characterized stream insects (Tebo and Hassler 1961) and trout in southern Appalachian streams (Tebo and Hassler 1963). Funding from the International Biological Program advanced CWT investigations of stream responses to forest management (IBP, Woodall and Wallace 1972, 1975, Webster and Patten 1979), and this research portfolio morphed into the LTER studies of how streams responded to the 1976 logging of WS 7 (summarized by Webster et al. 1983, 1992, Meyer et al. 2014, Wallace and Ely 2014, Swank and Webster 2014) discussed above.
Watershed-stream connection studies were broadened to include studies of other land uses including agriculture and development occurring within the upper Little Tennessee River basin (McTammany et al. 2007, 2008, Hagen et al. 2006, 2010, Webster et al. 2012, 2014, 2018, Burcher et al. 2007, Harding et al. 1998, Scott et al. 2002, Jackson et al. 2017). These studies have shown that relatively small amounts of forest conversion to small farms and rural residences results in substantial increases in stream DIN, specific conductivity, and sediment concentrations as well as summer stream temperatures. Other regional stream studies investigated how land use changes are reflected in biological changes along the Little Tennessee River, from its headwaters to where it is dammed up to form Fontana Reservoir (Neatrour et al. 2004, McTammany et al. 2003, Grubaugh et al 1996, Rosi-Marshall and Wallace 2002, Frisch et al. 2016, Cecala et al. 2018). Regional studies of the distribution of fish species indicates that exurbanization has favored cosmopolitan fish species over cold-water highland endemic species (e.g. Scott and Helfman 1999, Kirsch and Peterson 2014). Insect defoliation and loss of forest tree species by invasive insects have also been shown to impact Coweeta streams (Swank et al. 1981, Webster et al. 2012).
CWT studies were fundamental to the expansion of knowledge of the importance of wood to the ecology of forested streams. At Coweeta the role of wood in steams has been investigated in a variety of both observational and experimental studies including wood removal and wood replacement with PVC, and these studies examined interactions between wood, nutrient concentrations, basal resources, and invertebrate communities (Wallace et al. 2000, 2001, Webster et al. Eggert and Wallace 2007, Golladay and Webster 1988, Hedman et al. 1986, McTammany et al. 2008, Tank and Webster 1998, Tank et al 1998, Webster et al. 2014, Wallace et al. 1995, Webster et al. 1999, Webster et al. 1994)
CWT researchers conducted a suite of innovative whole stream experiments to elucidate stream responses to specific drivers of stream ecosystems. The elimination of stream insects by insecticide treatment demonstrated the importance of these insect as a connection between forests and the downstream transport of organic matter (Wallace et al. 1982, 1991, Cuffney et al. 1984, 1990). Covering a small stream with netting to sever the leaffall forest-stream connection clearly demonstrated the importance of allochthonous litter to stream insects (e.g., Wallace et al. 1997, 1999, 2015) and other aspects of stream ecosystem dynamics (Meyer et al. 1998 , Webster et al. 2001). Stream-reach scale nutrient addition studies have shown substantial interactions among nutrient concentrations, stream temperatures, and leaf litter composition in the breakdown of in-stream organic matter, and that higher nutrient levels accelerate leaf litter breakdown creating resource limitations in late summer and early fall (e.g., Benstead et al. 2009, Cross et al. 2006, Gulis et al. 2004, Manning et al. 2015, Suberkropp et al. 2010, Rosemond et al. 2015).
CWT researchers have advanced understanding of how individual fishes and species select microhabitats in a patchy stream environment to optimize metabological energetics (Petty and Grossman 1996, 2004 2009; Thompson et al. 2001). Long-term studies of fish assemblages along a longitudinal gradient have elucidated how environmental variation and density dependence interact to affect fish population dynamics (Grossman et al. 1998, 2006, 2016).
The nutrient spiraling concept, fundamental to all modern investigations of stream biogeochemistry, was developed at Coweeta to understand the behavior of the headwater streams used in the WS7 clearcut experiment (Webster 1975, Webster and Patten 1979). Subsequent investigations at Coweeta and elsewhere demonstrated that headwater streams were nutrient cycling hotspots. CWT investigators helped highlight the importance of headwater streams (Meyer and Wallace 2001, Meyer et al 2007)
Computational models of stream processes proved to be useful for synthesizing our understanding of Coweeta streams and watersheds (Webster 1983, Webster 2007, Webster et al. 2009, Webster et al. 2016, Lin et al. 2015). These models have also provided a mechanism to generate predictions of stream behavior in response to changes to the watershed-stream connections; predictions that can subsequently be tested by observational and experimental studies. In addition, Coweeta studies have contributed significantly to intersite comparisons of stream processes across a wide array of watershed vegetation (e.g., Webster and Meyer 1997, Mulholland et al. 1997, Peterson et al. 2001, Webster et al. 2003, Mulholland et al. 2008, Tank et al. 2018, Valett et al. 2008, Thomas et al. 2001, Pringle???)
River and Stream Channels
CWT researchers expanded our knowledge of how riparian vegetation along affects channel morphology, invertebrate and fish communities, amphibian movement, and stream temperatures. Moving down an Appalachian stream network, the system shifts from steep, often colluvial, headwater streams with small floodplains to flatter streams with larger floodplains. This topographic shift is usually accompanied by a shift in riparian vegetation from dense forest to either pasture, grass, or shrub vegetation. Dleigh, channel width
Widespread settlement by non-indigenous people during the late 19th and early 20th centuries motivated timber harvest and mechanized hillside cultivation that resulted in excessive regional soil erosion (Glenn, 1911). Ensuing post-settlement floodplain accretion rates have been an order of magnitude faster than pre-settlement rates (Figure 3), at 1-10 mm yr-1 versus 0.01-0.10 mm yr-1, respectively (Leigh, 2016). Consequently, alluvial bottomlands are draped by a distinctive stratum of post-settlement alluvium that buries the pre-settlement bottomlands. Rapid post-settlement accretion of tributary bottomlands ultimately transitioned to incision, terracing, and new floodplain formation following reforestation after 1930 that reduced upland sediment yields. However, the bottomlands of main stem rivers (e.g. Upper Little Tennessee and Tuckaseegee Rivers) continue to accrete bottomland sediment at rapid rates in accord with the "distributed sediment budget" concept of Trimble (1994). That is because tributary stream bank erosion and erosive new home and road development generates sediment that funnels through the low-order streams and accretes along the main stems.
Pre-settlement stream channels had lower sediment yields, greater sinuousity, gentler gradients, and smaller cross-sectionscompared to modern streams. Historically, channels were mechanically straightened (channelized), which inherently steepened the gradients, and the cross-sections grew larger to accommodate greater flood runoff from uplands with lowered infiltration rates caused by deforestation, crop cultivation, pastures, and home development. Evidence of pre-settlement physical conditions of rivers and streams in the Blue Ridge Mountains is provided by meandering paleochannels (meander scars), alluvial stratigraphy, and the earliest historical accounts by explorers. Pre-settlement streams with catchments larger than about 20 km2 clearly had single-channel meandering planforms as indicated by infilled meander scars preserved on alluvial bottomlands. Meander scars are visible on aerial imagery, but they are not very common because rapid post-settlement sedimentation (Leigh, 2016) covered and levelled much of the bottomlands, thereby obscuring the pre-settlement land surface and bottomland topography. Radiocarbon dates indicate meandering paleochannels range in age from several hundred to several thousand years, essentially spanning the entire Holocene (Leigh 2010, 2016). Although Walter and Merritts (2008) indicated that pre-settlement streams in the eastern United States exhibited “anabranching channels and riverine wetlands” this is not true for >20 km2 catchments in the Blue Ridge. In addition to meander scars, stratigraphic investigations reveal classic bedload/lateral-accretion/vertical-accretion facies assemblages that are characteristic of meandering streams (Price and Leigh 2006; Leigh and Webb, 2006; Leigh, 2010; Wang and Leigh, 2012; Wang and Leigh, 2016; Leigh, 2016), and in many cases the pre-settlement sediments are brown in color (oxidized), instead of grey (reduced) as expected for wetland soils. During his 1775 exploration down through the Upper Little Tennessee River valley near present-day Franklin, NC, William Bartram (1791) comments in many different passages (p. 345-358) on the meandering pattern of tributaries and the main stem of the Little Tennessee River. Interestingly, he also notes widespread meadows, pastures, cultivated fields of Native Americans, and wild strawberry fields on the bottomlands (rather than woodlands).
The conversion of forests to lawns and pastures has increased overland flow during storms and reduced low flows. Forested soils tend to have high porosity and low bulk density, and therefore infiltration rates in forested soils are so high that they are rarely exceeded by precipitation rates. Lawn and pasture soils are much denser, with lower porosities (Price et al. 2010). Thus, surface infiltration rates of lawns and pastures are about 1/10th those found in forested soils and are commonly exceeded by precipitation rates (Price et al. 2010). Soil compaction due to forest conversion also appears to reduce overall hydrologic storage in these watersheds, with the effect that late summer and fall low flows are lower in watersheds with less forest cover (Price et al. 2011).
While these significant changes in the hydrology of individual watersheds are important to the transport of pollutants and the maintenance of late summer stream habitat, streamflow changes due to rural development are small relative to the high degree of natural hydrologic variability in basins of the southern Appalachians owing to the high variability in climate and topography. For example, because of regional rainfall variability, unit-area peakflows and annual yields in the most urban stream in the region (Crawford Branch in Franklin, NC) are still substantially lower than unit-area peakflows in the fully forested Ball Creek which is much higher, steeper, and wetter (Jackson et al. 2017).
Because grasses have denser shallow root networks, the conversion of forested riparian zones to grass and pasture streambanks causes the active width and bankfull width of stream channels to become narrower for streams less than 20 m wide (Jackson et al. 2015, Leigh 2010, Faustini et al. 2009, Anderson et al. 2004, Hession et al. 2003). This reduction in stream width relative to forested riparian conditions is pronounced, such that active channel widths for grass-banked streams are generally about 1/3rd as wide as forested streams for the same basin area and valley slope (Jackson et al. 2015, Leigh 2010). This translates directly into a loss of benthic habitat area. Furthermore, forested streams have greater abundance of wood pieces in the channel, and this wood strongly affects habitat complexity by creating jams and steps in channels (Jackson et al. 2015, Jensen et al. 2014). Forested streams therefore feature much greater diversity of channel depths and velocities and larger numbers of flow obstructions (Jackson et al. 2015). Streams without any forested buffer have almost no in-channel wood. This isn’t just because of low recruitment, it is also because of active wood removal by stream-adjacent landowners who think the wood looks messy and who are concerned that it will block bridges and culverts during floods (Evans and Jensen-Ryan 2017). In summary, conversion of riparian vegetation from forests to grass or pasture has reduced benthic habitat area and aquatic habitat complexity. These space-for-time stream surveys also indicated that even modest riparian forest restoration could improve stream habitat. Narrow forest buffers of only 1 m width (trees on the streambank) were associated with wider channels and increased complexity relative to grass and pasture streambanks (Jackson et al. 2015). Streams with forest buffers 3-10m wide exhibit channel widths and habitat conditions intermediate between fully forested riparian zones and narrow buffers.
Forested streams in this region rarely experience summer water temperatures above 18oC due to high canopy cover and extensive shade (Swift and Messer 1971; Jackson et al. 2017; Long et al. 2014), but streams in agricultural and residential valleys often record summer temperatures that are 4-5oC warmer, in the range of 22-23oC (Jackson et al. 2017, Coats and Jackson 2020). These are temperatures that are very stressful to salamanders and trout adapted for cool mountain streams. Furthermore, streams in basins with mountainside housing developments have warmer temperatures than streams in basins with only valley development (Jackson et al. 2017). Small stream temperatures are very sensitive to canopy cover. Even understory rhododendron removal, leaving an intact overstory with greater than 82% canopy cover, still leads to 1-3oC increases in summer stream temperatures (Raulerson et al. 2020). Longitudinal surveys of summer stream temperatures above, within, and downstream of riparian canopy openings have revealed that smaller streams are more sensitive to changes in canopy cover, and these small streams can cool down rapidly after returning to forested riparian conditions (Coats and Jackson, 2020).
Land Use Legacies
Historic activities have imparted persistent legacies that affect contemporary ecological patterns and processes. Coweeta LTER researchers have studied how differences in land-use history affect the spatial distribution of mineral soil elements and nitrogen (N) cycling, microbial community composition and function, and the suitability of sites for plant establishment and survival. Although comparisons of averaged values rarely indicated that historical land use had an enduring effect on mineral soil nutrient pools or N cycling rates, differences in variance suggested that former human activities continue to influence nutrient distributions by altering their spatial heterogeneity. Patterns differed by element, but generally variance of soil carbon, N and calcium decreased and variance of soil phosphorus, potassium and magnesium increased with past land use intensity. Soil microbial analyses showed that microbial communities in formerly farmed stands had a higher relative abundance of markers for Gram negative bacteria and a lower abundance of markers for fungi compared with previously logged and reference stands. Net N cycling rates were negatively correlated with fungi in both farmed and reference stands, suggesting that historic land use can alter microbial communities in ways that may influence the processes they mediate. Geostatistical analysis showed that such differences in nutrient availability were an important factor in determining plant heterogeneity in reference stands, but not in previously altered stands. Species dispersal and reproductive syndromes also explained variability in the spatial heterogeneity of plants. Short-distance dispersal reduced plant heterogeneity regardless of past land use. In contrast, vegetative reproduction reduced heterogeneity only in reference stands. Plant biomass allocation patterns and growth rates varied considerably with past land use, but were associated only in historically unaltered stands. These results suggested that competition for light and nutrients may be reduced in areas with a history of human activity. Collectively, this research indicates that human activities have an enduring influence on the soil properties and compositions of soil microbial and understory plant communities.
Humans and the Landscape
CWT was one of only three LTER sites with an explicit mandate to include social science research in its portfolio and broadly investigate human/ecosystem interactions. Our research therefore expanded from an analysis of ecological dynamics in the uninhabited watershed managed by the US Forest Service at the Coweeta Hydrologic Laboratory to a consideration of socio-ecological processes across Southwestern North Carolina and even the surrounding Piedmont megapolitan archipelago. While the LTER remained predominantly an institution for natural and physical scientists, a number of social scientists were incorporated into the scientific team.
The Piedmont megapolitan region surrounding the southern Appalachian mountains provided a scale appropriate to integrate social and ecological research of the LTER Project, focusing on how exurbanization establishes regional patterns of urban metabolism (Gragson and Grove 2006). In southern Appalachia, Gragson and Bolstad (2006) tracked land use changes as seen through Coweeta LTER research. Much of this research shows increasing urbanization of these areas, especially in the last several decades, and forecasts significantly more urban growth until 2030 (e.g., Wear and Bolstad 1998). Other relevant Coweeta LTER research highlights the settlement patterns (Kirk, Bolstad and Manson 2012), bird populations (Lumpkin, Pearson, and Turner 2012), water quality issues (e.g. Webster et al. 2012, Jackson et al. 2017, Webster et al. 2018), channel responses to riparian management (Jackson et al. 2015), and aquatic organism populations (Scott 2006, Kirsch and Peterson 2014, Frisch et al. 2016, Cecala et al. 2018) intimately related to exurbanization in southern Appalachia. The exurban-themed research at the Coweeta LTER complemented other urban ecological research from the greater network. The Central Arizona Project and the Baltimore Ecosystem Study are two fully urban LTER sites studying urban ecology (e.g., Pickett and Cadenasso 2006; Childers et al. 2011).
Coweeta LTER researchers have augmented the 80+ year record of climate and hydrology from the Coweeta Hydrologic Laboratory with social, economic, and land-use records for the last 200+ years and paleoenvironmental records for the last 10,000 years. With these records and the reconstructions based on them, Coweeta LTER researchers are beginning to resolve the relative magnitude of climatic-versus-human-induced change and identify major turning points in the frequency of disturbance across the Holocene epoch in southern Appalachia.
Focus of New Research at Time of LTER termination
As discussed above, forest composition has changed substantially since the loss of Chestnut, and the forest is becoming mesophytic over time, with many biogeochemical and ecological implications. There are at least four, non-exclusive, reasons for this shift:
- Successional patterns following American chestnut loss.
- Enhanced biological nitrogen fixation after forest disturbance favors maples and tulip poplar.
- Fire exclusion has favored maples and tulip poplar.
- Shifting climate region: Greater precipitation over the last 100 years compared to the prior 500 years.
Improving our understanding of observed forest changes will continue to be a focus of research at the USFS Coweeta hydrologic laboratory following the end of LTER operations. Toward this end, the USFS is conducting a Paired Watershed Forest Composition Manipulation to move the treatment watershed (WS 31) toward an open OH forest using selective cutting, herbicides, and prescribed burns (standard silvicultural practices) following two years of pre-treatment monitoring. We will cut all understory evergreen shrubs (rhododendron and mountain laurel, Kalmia latifolia), herbicide stumps, allow material to dry so that fire can carry, and implement a prescribed burn with moderate intensity over the entire watershed. A moderately intense fire will consume the downed shrub layer, damage or kill some of the dominant tree species with thin bark (e.g., red maple, Bova and Dickinson 2005, Hengst and Dawson, 1994), and promote a more open canopy allowing oak and hickory species to regenerate after each scheduled burn (Arthur et al. 2015). Midstory competitors will be manually stem injected with herbicide as needed to allow oak speciesto recruit into the canopy (Loftis 2004). We will implement a prescribed burn 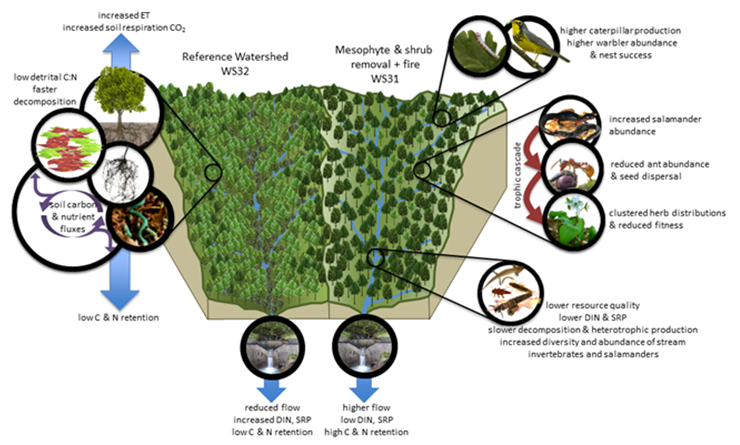 on a three-yr return interval, with the desired future condition of a moderately open woodland dominated by oak and hickory tree species with no evergreen shrub layer (Arthur et al. 2015). At the watershed-scale, we expect that a shift toward a more open and water-use-efficient OH forest will increase streamflow, produce more recalcitrant and lower quality leaf litter, and decrease stream nitrogen concentrations, with attendant effects on stream and forest animals. This watershed-scale experiment allows us to examine aspects of biogeochemical cycling, water and nutrients fluxes, and habitat responses that are not possible at the plot-scale.
on a three-yr return interval, with the desired future condition of a moderately open woodland dominated by oak and hickory tree species with no evergreen shrub layer (Arthur et al. 2015). At the watershed-scale, we expect that a shift toward a more open and water-use-efficient OH forest will increase streamflow, produce more recalcitrant and lower quality leaf litter, and decrease stream nitrogen concentrations, with attendant effects on stream and forest animals. This watershed-scale experiment allows us to examine aspects of biogeochemical cycling, water and nutrients fluxes, and habitat responses that are not possible at the plot-scale.
The treatment watershed (WS31) is an east-facing, mid-elevation (869–1141m) watershed, 37.8 ha in area, with streamflow and chemistry recorded sub-hourly or weekly, respectively, since 1981. The reference watershed (W32) is similar in aspect (east-facing), elevation (920–1236m) and area (41.3 ha); and it also has a longer record depth (streamflow since 1941 and chemistry since 1976). QA/QC protocols and equipment for all measurements and data acquisition will follow established protocols (Laseter et al. 2012) except where noted below. This experiment allows us to ask the question, how would an oak-hickory future forest affect hydrological and biogeochemical fluxes at the watershed-scale?
The above is just a sampling of CWT research and findings. As of June 2020, the CWT publication database included ~1496 journal articles, books, and book chapters and 318 graduate student theses and dissertations.



